Сборник идеальных эссе по обществознанию
Содержание:
Сравнительно-анатомические доказательства эволюции
Все позвоночные животные имеют двустороннюю симметрию, полость тела, позвоночник, череп, две пары конечностей. Сердце у всех позвоночных животных расположено на брюшной стороне, а нервная система — на спинной, она состоит из головного и спинного мозга. Единство плана строения в каждом типе свидетельствует о единстве его происхождения.

Двусторонняя симметрия — левая половина тела является отражением правой
Гомологичные органы
После выхода в свет работ Дарвина сравнительная анатомия получила толчок к развитию и в свою очередь внесла значительный вклад в развитие дарвинизма.
Большую роль сыграло установление гомологичности органов. Гомологичные органы могут выполнять различные функции и в связи с этим несколько разниться в строении, но построены по одному плану и развиваются из одних и тех же зародышевых зачатков.
Таковы передние конечности всех позвоночных: нога кролика, крыло летучей мыши, ласт тюленя, рука человека. Скелет каждого из этих органов имеет плечо, предплечье, состоящее из двух костей, кости запястья, пястья и фаланги пальцев. То же относится и к задним конечностям. Было обнаружено, что млечные железы гомологичны потовым, челюсти ракообразных — их конечностям, волосы млекопитающих — перьям птиц и чешуе рептилий, зубы млекопитающих — чешуе акул, части цветка (пестик, тычинки, лепестки) — листьям и т.д.
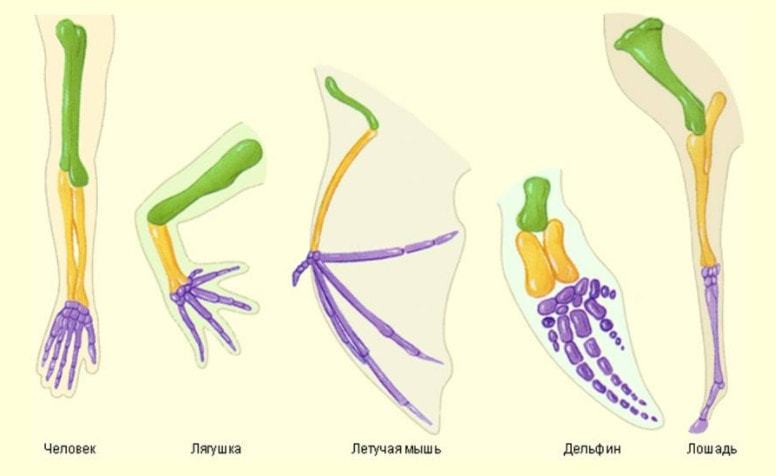 Примеры гомологичных органов
Примеры гомологичных органов
В отличие от гомологичных, аналогичные органы могут быть сходны по строению, так как выполняют однородные функции, но не имеют общего плана строения общего происхождения. Примерами их могут быть крыло насекомого и крыло птицы, жабры ракообразных и жабры рыбы. У растений аналогичными являются колючки кактуса (видоизмененные листья) и шипы розы (выросты кожицы). Для установления родственных связей между организмами они роли не играют.
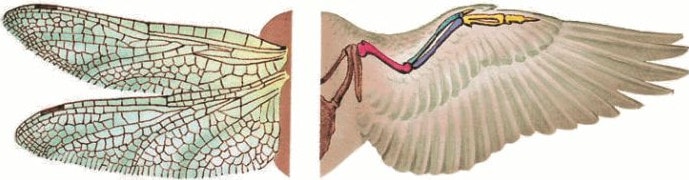 Пример аналогичных органов
Пример аналогичных органов
Атавизмы и рудименты
Для доказательства эволюции имеют значение атавистические органы, которые были присущи далеким предкам и в норме не встречающиеся у современных организмов. Естественно, что такие признаки говорят о филогенетическом родстве. Примерами атавизма служат появление боковых пальцев у лошади, полосатость у домашних свиней; шейная фистула (образование, гомологичное жаберным щелям у низших хордовых), хвостовой придаток, обильная волосатость всего тела у человека.
Рудиментарными называются органы, утратившие свою функцию, но сохраняющиеся у взрослых животных. Обычно они остаются в зачаточном состоянии. Рудиментарными являются остатки тазовых костей у безногой ящерицы желтопузика и у китообразных. Они служат доказательством происхождения этих животных от предков, имевших развитые конечности. У человека рудиментарными органами являются:
- Копчик — остаток хвостовых позвонков;
- зачаточные ушные мышцы свидетельствующие о том, что предки человека обладали подвижной ушной раковиной.
На корневищах папоротника, пырея, ландыша можно обнаружить чешуйки — рудименты листьев.
Сравнительно-анатомические исследования современных прогрессивных и примитивных форм позволяют обнаружить переходные формы. Морское животное баланоглосс сочетает в себе признаки животных типа иглокожих и типа хордовых. Ланцетник имеет ряд признаков, сближающих его с одной стороны с иглокожими и полухордовыми (баланоглосс), а с другой стороны с позвоночными, с которыми он относится к одному типу хордовых.
 Баланоглосс
Баланоглосс
Среди современных млекопитающих существуют однопроходные (имеющие клоаку и при размножении откладывающие яйца, как пресмыкающиеся), сумчатые и плацентарные. Сравнение их указывает, что млекопитающие находятся в родстве с пресмыкающимися и что эволюция млекопитающих шла от животных, откладывающих яйца, к живородящим формам с еще недоразвитой плацентой и, наконец, к животным, рождающим уже хорошо сформированных детенышей.
Примеры среди животных
Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб.
Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное.
Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобный современных птиц, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободные пальцы на крыльях, подвийноувигнути позвонки, длинный хвост с 20-21 позвонка, кости могло не пневматизовани, грудная кость без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса.
Большое количество ископаемых форм звероподобных пресмыкающихся (синапсиды, терапсид, пеликозавров, различных динозавровых и др.), Найденных во многих районах земного шара, существовали в юрский и меловой периоды, сочетающие признаки пресмыкающихся и млекопитающих, раскрывают возможные направления и способы становления различных групп четвероногих, в частности млекопитающих. Например, звероподобный пресмыкающееся из группы терапсид — лиценопс (Lycaenops)
по развитию костей ротовой полости, дифференцированием зубов на клыки, резцы, по резцовые зубы и рядом других признаков строения тела напоминает хищных млекопитающих, хотя по другим признакам и образом жизни это были настоящие пресмыкающиеся.
Одной из форм, сохранилась в ископаемом состоянии является амбулоцетус Ambulocetus natans
(«ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторинноводнимы формами. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа должна частично редуцированную шерсть. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде.
Переходные формы и таксономия
Эволюционная таксономия оставалась доминирующей формой таксономии течение ХХ в. Выделение таксонов базируется на различных признаках, вследствие чего таксоны изображают в виде ветвей разветвленного эволюционного дерева. Переходные формы рассматриваются как «падающие» между различными группами в плане анатомии, они смесь характеристик от внутренней и внешней клади, что недавно разделилась.
С развитием кладистики в 1990-х гг. Взаимосвязи обычно изображают в виде кладограмы, иллюстрирующую дихотомическое ветвление эволюционных линий. Поэтому в кладистици переходные формы рассматриваются как более ранние ветви дерева, где еще не развились не все черты, характерные для ранее известных потомков на этой ветке. Такие ранние представители группы обычно называют основным таксоном (англ. Basal taxa)
или сестринским таксоном (англ. Sister taxa),
в зависимости от того, принадлежит ли ископаемый организм к данной клади или нет.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами . Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.
Недостающие звенья
| 1850 | |
| 1900 |
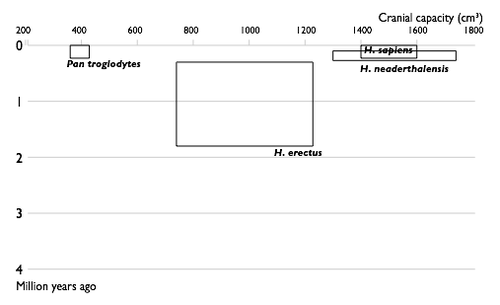 |
| 1950 |
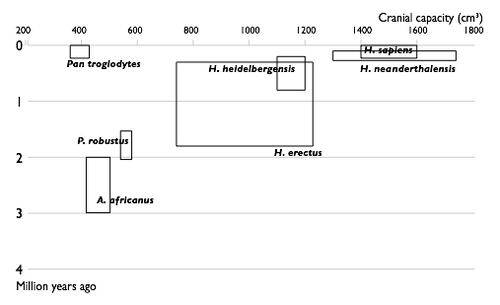 |
| 2002 |
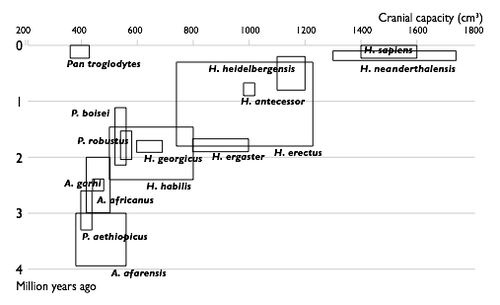 |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа , и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. |
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований . Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден Тиктаалик — ещё одно переходное звено между рыбами и земноводными)
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации , то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород . Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия .
Примеры среди растений
Первые наземные растения из класса риниопсид, семей риниевих и псилофитовых, живших в силуре — девоне, сочетали признаки зеленых водорослей и примитивных форм высших растений. Их тело было безлистным, цилиндрический осевой орган — телом в верхней части дихотомически разветвленным на верхушках с спорангиями. Функцию минерального питания риниопсид выполняли ризоиды.
Ископаемые формы семенных папоротников, которые процветали в конце девона, сочетают в себе признаки папоротников и голосеменных. Они образовывали не только споры (как папоротники), но и семена (как семенах растения). Проводящая ткань их стеблей по строению напоминает древесину голосеменных (саговников).
Другой предшественник семенных растений был идентифицирован из отложений среднего девона. Рункария (Runcaria heinzelinii)
существовала около 20 млн лет назад. Это была небольшая растение с радиальной симметрией; имела спорангий, окруженный интегументом и плюской. Рункария демонстрирует путь эволюции растений от споровых к семенных.








