Динозавры
Содержание:
Цератопсы,
или рогатые динозавры, появились среди гигантских рептилий последними. Их череп оканчивался высоким и узким беззубым клювом, а сзади разрастался в широкий костный воротник, который нависал над шеей примерно настолько же, насколько простиралась вперед остальная часть головы. Известно несколько вариантов «рогатости» этих рептилий. Некоторые из них практически безроги, другие несли лишь один рог – на рыле, а, скажем, у рода Triceratops их было три – по одному на рыле и над глазами. Традиционно считается, что эти рога использовались в схватках с хищниками. Однако аналогия с ныне живущими рогатыми животными, например буйволом, наводит на мысль об определенной роли таких структур в распознавании полового партнера и других формах социального поведения, особенно связанных с доминированием и спариванием.
Задние ноги цератопсов длинные и тяжелые, а передние, хотя и не столь внушительные, для птицетазовых относительно крупные; лапы широкие и короткие. Туловище было толстым, а хвост массивным и коротким, приблизительно равным ему по длине. Панцирь, если не считать головного воротника, отсутствовал.
Цератопсы известны только из мела, однако они успели дать множество разнообразных морфологических типов. Protoceratops из Монголии – наиболее известная мелкая примитивная форма. На серийном материале, собранном экспедицией Американского музея естественной истории, изучен процесс индивидуального роста этих ящеров, в частности развития воротника или гребня на заднем крае их черепа. Сначала его разрастание увеличивало площадь крепления мышц, приводящих в движение нижнюю челюсть. На более поздних стадиях, а также по мере специализации цератопсов воротник, вероятно, принимал на себя функцию защиты шеи как наиболее уязвимой области тела. С учетом данных, указывающих на возможную теплокровность этих ящеров, т.е. их способность поддерживать постоянную температуру тела, можно предположить, что воротник участвовал и в ее контроле, при избытке тепла рассеивая его, а при недостатке – улавливая из внешней среды.
Особенно много разнообразных цератопсов обнаружено в верхнемеловых отложениях Азии и Северной Америки. Больше других известен Triceratops длиной ок. 6 м. Его череп нес огромный воротник и три хорошо развитых рога. Ноздри были очень велики, верхняя челюсть завершалась мощным клювом, щечные зубы развивались слабо. Несколько меньший Monoclonius отличался присутствием всего одного рога на морде. У рода Styracosaurus тоже был лишь один носовой рог, зато его воротник окаймляли длинные шипы.
Цератопсы относятся к наиболее многочисленным динозаврам конца мелового периода. Вероятно, они были хорошо адаптированы к жизни на открытых пространствах и питанию цветковыми растениями. Несмотря на обилие этих ящеров, прежде всего из рода Triceratops, они не пережили перехода от мезозоя к кайнозою, и их вымирание ознаменовало конец эпохи динозавров.
В начале 1920-х годов в нижнемеловых отложениях монгольской пустыни Гоби обнаружены гнезда с яйцами Protoceratops. Со времени этого открытия было найдено много других гнезд динозавров. Прекрасно сохранившиеся захоронения такого типа раскапывают в Монголии. Они содержат целые кладки яиц утконосых динозавров с невылупившимися эмбрионами и скелеты их детенышей. Там же найдены гнезда мелких двуногих орнитопод и неизвестных крупных животных.
Как показали эти находки, уход за гнездом и забота о потомстве у утконосых динозавров были сложнее, чем у современных крокодилов, которые обнаруживают лишь зачатки родительского поведения. Такие особенности, как строительство гнезда, признаки заботы о потомстве вместе с возможной теплокровностью, широким разнообразием типов динозавров и освоенных ими местообитаний, а также свидетельствами их стадного поведения, позволяют говорить о сложной социальной структуре сообществ этих мезозойских рептилий, развивавшейся на протяжении 100 млн. лет их господства. Возможно, по уровню поведенческой организации они приближались к современным птицам или млекопитающим.
Орнитоподы.
Среди древнейших остатков птицетазовых изредка встречаются «птиценогие» ящеры, датируемые поздним триасом или самым началом юры. Однако немногие хорошо сохранившиеся образцы и некоторые фрагментарные остатки дают достаточно ясное представление об этих животных. Лучше других известен Heterodontosaurus, двуногое существо длиной ок. 0,9 м, а также род Fabrosaurus. Судя по ним, в конце триаса уже существовало по крайней мере два четко обособленных семейства орнитопод, следовательно, вся эта группа оформилась не позднее среднего триаса. Некоторые архаичные ее представители, т.н. эволюционные реликты, встречаются и в отложениях мелового возраста. Таков, например, род Hypsilophodon из нижнего мела. Это было маленькое быстро бегающее двуногое животное, примитивность которого выражалась в сохранении передних зубов на верхней челюсти. Хвост его длинный и тонкий, а вытянутые пальцы задних ног, вероятно, хорошо гнулись. Сравнительно примитивная юрская форма Camptosaurus дает представление о промежуточной эволюционной стадии между ранними и более специализированными орнитоподами. Это был тяжелый ящер с довольно крупными передними конечностями, позволявшими предположить, что передвигался он в основном на 4 ногах. Зубы и челюсти хорошо приспособлены для питания растительным кормом, а беззубый клюв служил, вероятно, для скусывания сочных побегов.
Более специализированны «утконосые» орнитоподы, называемые траходонтами или гадрозаврами. У них был плоский клюв без передних зубов. Однако в щечной области челюсти несли мощные перетирающие зубы, хорошо приспособленные к питанию грубой растительной пищей. Утконосые динозавры были крупными, тяжеловесными животными. Длина их тела в среднем превосходила 9 м. Задние конечности были массивными, а передние хотя и более короткими, но хорошо развитыми. Обычно они передвигались на двух ногах, но могли становиться и на четвереньки.
Среди утконосых динозавров обнаружены колоссальные различия в строении черепа, касающиеся в первую очередь окружающих ноздри костей. У некоторых они образуют причудливые вздутия в носовой области, у других полые куполообразные гребни или длинные выступающие позади головы трубчатые структуры.
По-видимому, в конце мелового периода утконосые динозавры были распространены почти по всему свету. В Северной Америке они лучше всего известны из Канады, где представлены как консервативные формы, например Anatosaurus, так и специализированные типы вроде Lambeosaurus с высоким вздутым гребнем на голове. Их остатки встречаются главным образом в осадочных породах, сформировавшихся на дне рек, озер и болот. Вероятно, эти животные вели амфибиотический образ жизни и хорошо плавали. Известны также их мумифицированные остатки с сохранившимся рельефом кожи и очертаниями формы тела. Они показывают, что тело покрывали мелкие чешуи, панциря не было.
Прозавроподы.
К этой группе относятся триасовые представители завроподоморф. Наиболее примитивен среди них род Thecodontosaurus из позднего триаса Англии, вероятно представляющий собой сестринскую группу по отношению ко всем остальным прозавроподам (т.е. параллельную им ветвь эволюции). Этот динозавр был двуногим. Прозавроподы в конце триасового периода быстро эволюционировали. Они известны на всех континентах, кроме Австралии и Антарктиды, и составляют неоднородную группу, включающую приблизительно 17 родов из 7 семейств. Типичным представителем может служить Plateosaurus, достигавший в длину около 6 м. Задняя часть его тела была очень массивной, как у многих теропод, однако довольно хорошо развитые передние конечности, вероятно, позволяли передвигаться как на двух, так и на четырех лапах. Широкие стопы мало походили на птичьи. Длинная шея несла маленькую голову. Сравнительно слабые челюсти усажены лопатовидными или листовидными зубами, которые напоминают известные у некоторых настоящих завропод.
Анкилозавры
(панцирные динозавры) составляют крайне интересную, но недостаточно изученную группу птицетазовых. Известны они прежде всего из меловых отложений Северной Америки, Европы, Азии и, возможно, Южной Америки. Их остатки немногочисленны, и эволюционная история остается неясной. Эти динозавры удачно названы «рептилиями-танками», их крупные представители достигали в длину 6 м. У типичных анкилозавров из родов Nodosaurus, Ankylosaurus и Palacoscincus панцирь устроен следующим образом. Чередующиеся ряды крупных и мелких многоугольных пластин формировали покрывающую все тело защитную мозаику, на ногах были шипы, на хвосте – панцирные кольца, в некоторых случаях с шипами, а на голове как бы сплошной костный шлем. Форма туловища тяготела к широкой и уплощенной, а держалось оно на коротких мощных конечностях. Находящиеся в Москве находки из Монголии позволяют проследить ряд форм, сменявших друг друга с нижнего мела до конца этого периода. Лучше всего изучен Syrmosaurus, внешним видом несколько напоминающий многократно увеличенную жабовидную ящерицу. Самым крупным был Ankylosaurus, достигавший в длину 10,7 м.
Большинство анкилозавров было медлительными фитофагами. Их слабые челюсти и почти отсутствующие у некоторых форм зубы указывают на питание мягкими частями растений. Об их происхождении почти ничего не известно, хотя некоторые формы из раннего мела Европы несколько примитивнее типичных и лучше изученных родов.
Тероподы
(звероногие) – это широкий спектр преимущественно двуногих форм, известных с позднего триаса до самого конца мелового периода. До недавнего времени их условно делили на две группы – целурозавров и карнозавров. Целурозавры – это двуногие животные мелких-средних размеров с длинными задними конечностями и маленькими, облегченными передними. Длинный хвост предположительно служил им балансиром при двуногой (бипедальной) локомоции. Находки показали, что целурозавры представляют собой не единый кладон, а несколько четко обособленных эволюционных линий. Долгое время их считали самыми примитивными тероподами, однако лидирует в этом смысле группа карнозавров, названная цератозаврами. Она представлена крупной юрской формой Ceratosaurus с небольшим рогом на носу, родом Coelophysis, известным по многим экземплярам из триаса штата Нью-Мексико, а также несколькими другими триасовыми родами из Северной Америки, Европы и Южной Африки.
Родственные связи различных групп целурозавров не вполне выяснены и по-разному интерпретируются специалистами. Особый интерес представляют их более эволюционно продвинутые формы, включая птиц. Среди них выделяются «страусоподобные динозавры», например Struthiomimus, Ornithomimus и Gallimimus. Это были бегающие тероподы средних-крупных размеров с облегченным телосложением. Передние конечности их длинные, но кисти не хватательные; задние ноги стройные, с компактными стопами; челюсти беззубые, морда тонкая и вытянутая. Только один род юрский, все прочие – меловые. Менее известны овирапторозавры, мелкие редкие двуногие формы длиной не более 2 м, также лишенные зубов. Однако их череп короткий и высокий, с сильно редуцированными лицевой частью и передними костями челюстей.
К наиболее интересным «некарнозаврам» относится группа т.н. дромеозаврид. Это двуногие существа мелких-средних размеров, известные из юры и мелового периода. Один из наиболее изученных их родов – Deinonychus из штата Монтана. Его четкий отличительный признак связан со строением стопы. Второй палец сильно увеличен, не служит опорой и оканчивается серповидным когтем, бывшим, вероятно, оружием нападения. Таким образом, стопа стала двупалой в противоположность обычной для динозавров трехпалости. Своеобразен также длинный и тонкий хвост, жесткий на протяжении примерно 3/4 своей длины, считая от конца.
К дромеозавридам близки редко встречающиеся троодонтиды, мелкие двуногие формы с самыми крупными среди всех динозавров относительными размерами мозга. Обе группы обладают многими признаками птиц.
Плотоядные карнозавры, например юрский Allosaurus и Tyrannosaurus из мелового периода, хорошо известны и представлены сравнительно немногими родами. Предшественником крупных юрских и меловых хищников был триасовый Palaeosaurus, довольно тяжелая двуногая плотоядная форма длиной ок. 2,7 м. В течение юрского периода доминировали гораздо более массивные ящеры, такие, как Allosaurus и Megalosaurus. Длина первого ок. 9 м. Его передние конечности сильно редуцированы, но еще достаточно крупные, чтобы функционировать. В противоположность ему у еще более крупных меловых форм, таких как Tyrannosaurus, передние конечности превратились в миниатюрные бездействующие придатки. Достигавший 15 м Tyrannosaurus был самым крупным когда-либо существовавшим наземным хищником. У этих гигантов голова относительно крупная. Вероятно, для снижения веса их череп приобрел крупные окна, не проиграв в прочности. Массивные челюсти усажены длинными, кинжалообразными зубами, прекрасно подходящими для разрывания жертвы. Похоже, что в течение юрского и мелового периодов эти хищники охотились на крупных растительноядных динозавров и, возможно, других рептилий соответствующих размеров. Главными их жертвами в юре были завроподы и стегозавры. В меловом периоде важным источником корма для этих крупных хищников, видимо, оставались некоторые завроподы, к которым добавились новые птицетазовые формы, в частности цератопсы и утконосые динозавры.
ПТИЦЕТАЗОВЫЕ
Этот монофилетический отряд более разнообразен по составу, чем ящеротазовые. Все птицетазовые были строго растительноядными и в целом эволюционно более продвинутыми, чем рассмотренные выше динозавры. Специализация затронула как фундаментальные особенности их строения, например форму таза, ставшего похожим на птичий, так и адаптации, связанные с питанием, передвижением и защитой от врагов. Наибольшего процветания группа достигла в меловом периоде, что совпало с распространением цветковых растений
Впрочем, хотя птицетазовые и доминировали среди наземных рептилий в позднем мелу, важной частью фауны были в это время также ящеротазовые хищники, страусоподобные динозавры, некоторые завроподы и птицы
Традиционно выделяли 4 сравнительно слабо различающиеся по уровню организации группы птицетазовых. К орнитоподам («птиценогим») относятся как примитивные, так и более специализированные двуногие рептилии. Три другие группы образованы высоко специализированными растительноядными формами, передвигавшимися на четырех ногах. Это преимущественно юрские стегозавры (пластинчатые динозавры), теловые анкилозавры (панцирные динозавры) и цератопсы (рогатые динозавры). Кроме похожего на птичий таза, для них характерно присутствие в нижней челюсти беззубой предзубной кости, расположенной спереди от типичной для всех динозавров зубной, несущей зубы. Впрочем, на обеих челюстях передних зубов, как правило, не было, и оканчивались они клювом. Зубы находились только в щечной области и были преобразованы для перетирания и разрезания растительного корма. В отличие от завропод, у некоторых из этих групп зубной аппарат высокоспециализированный и довольно мощный. Общие особенности таза и челюстей свидетельствуют, что все птицетазовые, несмотря на их различия, произошли от общего предка.
В то же время, как показал кладистический анализ, традиционное разделение этого отряда на 4 группы неудовлетворительно. Стегозавров и анкилозавров теперь объединяют в таксон Thyreophora. В его рамках к наиболее архаичным, возможно, наряду со скорее всего двуногим Scutellosaurus, относятся сцелидозавриды, снабженные легким панцирем формы, передвигавшиеся на четырех ногах. Они появились приблизительно на рубеже триаса и юры. Однако еще примитивнее триасовые роды Lesothosaurus (Южная Африка), Technosaurus (Северная Америка) и Pisanosaurus (Южная Америка). Их родственные связи с остальными птицетазовыми пока не вполне выяснены. Орнитоподы, ранее считавшиеся наиболее примитивными из птицетазовых, по-видимому, эволюционно более продвинуты, чем древнейшие тиреофоры. Последними появились цератопсы, включавшие протоцератопсов и собственно цератопсов, иногда называемых неоцератопсами. «Попугаеклювые» пситтакозавры и купологоловые пахицефалозавры связаны общими предками с цератопсами.
ПРЕДКИ ДИНОЗАВРОВ
В начале триасового периода, перед появлением динозавров, на Земле существовала разнородная группа текодонтов – архозавровых рептилий мелких и средних размеров. Среди них выделялись легко сложенные и быстро бегающие двуногие формы, известные под названием псевдозухии («ложные крокодилы») и представленные, например, родом Saltoposuchus. Особенностями скелета они напоминали динозавров, но были менее специализированными. Текодонты опирались в основном на всю стопу (стопохождение), тогда как динозавры – только на пальцы (пальцехождение).
Предками их были, безусловно, какие-то двуногие текодонты. Таз последних примерно такой, как у ящеротазовых, к которым и могла вести от них прямая линия развития. Таз птичьего типа, по-видимому, возник у динозавров позже. У птиц он, возможно, развился независимо, поскольку они произошли от ящеротазовых. Обычно птиц выделяют в самостоятельный класс Aves, но иногда предлагают рассматривать их как одну из групп тероподовых динозавров, т.е. как оперенных теропод, в составе отряда Saurischia.
Непосредственные предки текодонтов точно не известны. Более 225 млн. лет назад, в пермском периоде палеозойской эры, среди примитивных рептилий выделилась группа, известная под названием «эозухии» («ранние крокодилы»). Внешне они напоминали ящериц длиной ок. 30 см. Позже из эозухий возникли ящерицы, змеи и новозеландская гаттерия. От них же могли произойти и текодонты, но этот вопрос остается открытым. Если они действительно потомки эозухий, эту группу следует считать предковой для всех ныне живущих рептилий, за исключением черепах. Последние произошли независимо, от еще более примитивных форм. Прямые предки черепах неизвестны, но, возможно, к ним близка мелкая зубастая рептилия Procolophon пермского возраста.
Об особенностях примитивных динозавров позволяют судить недавние находки позднетриасовых представителей этой группы в Аргентине, Бразилии и Южной Африке. Среди них хорошо сохранились череп и другие части скелета мелкого, легко сложенного животного, названного Eoraptor. Судя по этому, а также другим примитивным триасовым родам, например Herrerasaurus и Staurikosaurus, первые динозавры были двуногими. Они отнесены к тероподовым ящеротазовым, но набор диагностических признаков этой группы у них неполон, а их таксономическое положение остается спорным. Современником этих теропод был довольно плохо сохранившийся род Pisanosaurus из Аргентины. Он считается птицетазовым. Сходные, но несколько более поздние примитивные птицетазовые известны по находкам в Южной Африке. Очевидно, что разделение динозавров на ящеротазовых и птицетазовых произошло в скором времени после их возникновения из нединозавровых предков.
Когда в конце триаса на Земле появились динозавры, климат на значительной части суши, по-видимому, был довольно прохладным и сухим, способствовавшим эволюции быстроногих животных. Более ранние наземные позвоночные населяли преимущественно влажные низменности с обилием озер, рек и болот и достаточно высокими температурами. Ящеротазовые динозавры одними из первых приспособились к климатическим особенностям мезозоя. Однако они достигли своего расцвета в юрском периоде, когда на большей части Земли снова воцарились теплые и влажные условия. Из триасовых отложений известны как ящеротазовые, так и птицетазовые, но обильными в то время были только первые.
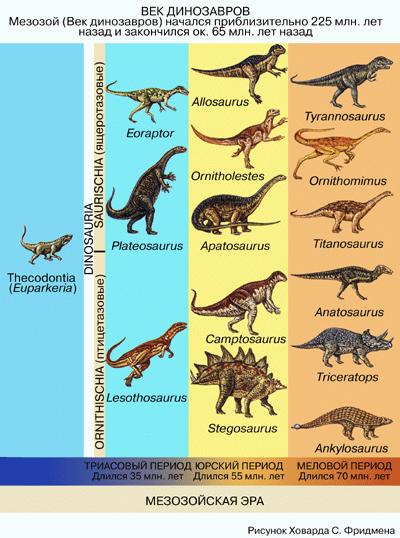



КОНЕЦ ЭРЫ ДИНОЗАВРОВ
В позднем мелу сложная экосистема, в которую входили динозавры, включала также цветковые растения, многие типы насекомых, ящериц, змей, черепах, крокодилов, а также летающих рептилий и птиц. В ней присутствовали и очень мелкие млекопитающие. С окончанием мелового периода вымирание динозавров резко изменило эту динамичную биоту.
Что касается динозавров, остаются нерешенными два особенно интересных вопроса. Почему столь многие их линии развивались в сторону увеличения размеров, и от чего добившиеся такого явного успеха формы вымерли? В обоих случаях можно сделать лишь некоторые предположения. Многие группы животных в ходе своей эволюции имели тенденцию к увеличению размеров. В общем, крупные размеры особей в популяции дают им преимущества перед остальными в смысле защиты от врагов, добывания корма, спаривания и т.д. Однако в эволюции динозавров такая тенденция привела к явным излишествам. Не вполне понятно, отчего так получилось. Вероятно, сыграли свою роль устойчивая температура и обилие растительного корма. За увеличением размеров фитофагов почти наверняка последовало укрупнение питавшихся ими хищников. Межвидовая и внутривидовая конкуренция и естественный отбор привели к своего рода порочному кругу, точнее, спирали, ведущей к гигантизму. Конечно, он присущ не только динозаврам и возникал также у рыб, морских рептилий, летающих ящеров и млекопитающих, среди которых появились, в частности, огромные киты.
Неясными остаются причины вымирания динозавров. В конце мезозоя изменились физические условия среды. Вероятно, они стали для динозавров менее благоприятными, чем ранее. Однако в прежние эпохи эти ящеры переживали гораздо более значительные смены условий, и некоторые относительно неспециализированные их формы были в состоянии адаптироваться к экологическим сдвигам такого масштаба, какой отражен в геологической летописи конца мелового периода.
К тому же важно учесть, что вымерли не только динозавры. Окончания мела не пережили некоторые другие наземные животные, ряд пресноводных и морских рыб, а также морские рептилии – плезиозавры и мозазавры
Вымирание, или серьезное сокращение разнообразия и численности, постигло также определенные группы морских беспозвоночных. Нет сомнений, что эволюционный упадок всех этих организмов каким-то образом обусловлен внешними событиями.
В большинстве современных исследований непосредственной причиной вымирания динозавров считается глобальная катастрофа. Интенсивно изучаются климатические последствия, к которым могло привести столкновение Земли с кометой или огромным астероидом. Во многих местах планеты на границе между меловыми и третичными породами залегает тонкий слой глины с намного превышающим норму содержанием редких металлов, включая иридий. Многие полагают, что такая высокая их концентрация при столь широком распространении может быть обусловлена только столкновением с космическим телом, например с гигантским метеоритом, планетоидом или кометой. Тектонические свидетельства колоссальной силы удара в конце мела недавно обнаружены в области Карибского бассейна. Однако другие ученые считают, что вымирание могло наступить в результате интенсивной вулканической деятельности, которая также зафиксирована геологической летописью в самом конце мелового периода.
Вместе с тем, даже если земная среда обитания действительно быстро изменилась вследствие одного или серии мощных катаклизмов, вопрос о влиянии этих экологических сдвигов на динозавров остается открытым. В конце мела быстро исчезли аммониты, процветавшая в мезозое группа головоногих моллюсков. В это же время резко изменились одноклеточные морские организмы фораминиферы. Однако к тому моменту уже произошло постепенное сокращение разнообразия и численности динозавров. Если тогда и и наступила глобальная катастрофа, это могло послужить причиной исчезновения лишь последних представителей их некогда могучего племени, если только оно не угасло еще раньше.
Таким образом, вполне вероятно, что в основе вымирания динозавров, как и других массовых вымираний, происходивших в истории Земли, лежит сложный комплекс эволюционных преобразований растительного и животного мира, а также физических событий, связанных с изменениями площади и очертаний суши и моря, горообразовательными процессами, распределением температур, ветров и океанических течений.
См. также ПТЕРОЗАВРЫ; ПРЕСМЫКАЮЩИЕСЯ; ПАЛЕОНТОЛОГИЯ; ГЕОЛОГИЯ.